(páginas 22-43 de De la Puente, J., Pérez-Tris, J. y Bermejo, A. (2003). Anuario Ornitológico de Madrid 2002. SEO-Monticola, Madrid).
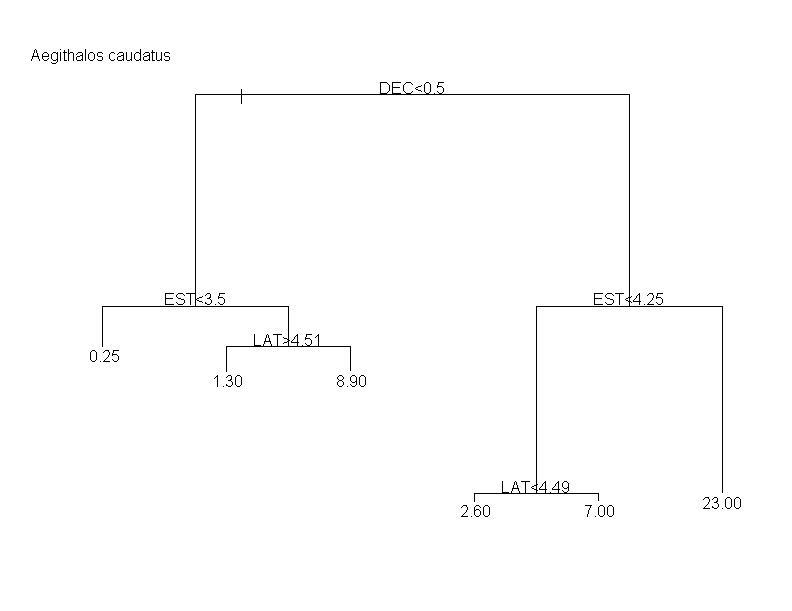
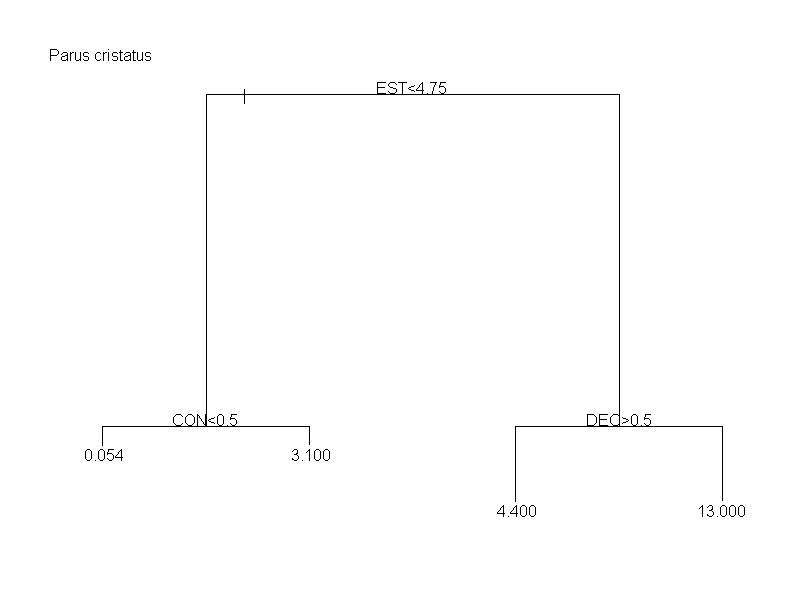
Los dendrogramas resultantes de los análisis de regresión en árbol constan de varios elementos. Los nodos o dicotomías, distinguen pares de posibilidades de abundancia o densidad para la especie, que le confieren su apariencia ramificada y que en cada caso se rigen por un criterio o condición de naturaleza discreta. Estos criterios son el valor que toma una variable descriptora para dividir las observaciones en dos subconjuntos tan diferentes entre sí como sea posible (de forma similar a los pasos de una clave de identificación de especies). Cada criterio se identifica con el nombre de la variable independiente a la que hace referencia y con una condición que, cuando es verdadera, se cumple para los elementos de la rama izquierda (ver más adelante). Las ramas resultantes de cada dicotomía, indican gráficamente la importancia de cada uno de los criterios en la descripción general de las preferencias de la especie. Las hojas o puntas terminales de los árboles, informan de la abundancia relativa en las cuadrículas del atlas (aves/20 horas) o densidad media en los censos (aves/30 ha) de todas las unidades muestrales agrupadas en ellas. Como guía ilustrativa se explican los patrones observados para el Mito (Aegithalos caudatus) y para el Herrerillo Capuchino (Parus cristatus), considerando tanto los árboles de regresión resultantes de los datos de abundancia del atlas de invernantes, como los de densidades en los 77 censos acumulados.
En A. caudatus el primer nodo o criterio que divide las abundancias estimadas por el atlas (fig) en dos grupos homogéneos está determinado por el porcentaje de suelo dedicado a uso agrícola. En la parte superior del árbol se indica "agrícola>0.36", lo que debe interpretarse como que las cuadrículas con un porcentaje superior al 36% de suelo agrícola se encuentran en las puntas terminales de la izquierda. En cambio, el primer criterio para P. cristatus (fig), que es "conífera <0,11", indicaría que las unidades muestrales que tienen menos del 11% de su superficie ocupada por masas de coníferas están a la izquierda del árbol. Para el caso de los datos de densidad procedentes de los 77 censos, el primer criterio que divide las densidades de A. caudatus (fig) obtenidas en dos grupos homogéneos está determinado por el criterio "deciduos<0.5". Esta es una de las variables de tipo binomial consideradas (sólo presentan dos posibles valores: ‘ausencia’=0 y ‘presencia’=1). Así, esta condición nos estaría indicando que los censos sin presencia de arbolado caducifolio (más tendentes a 0, como promedio), se acumulan en las puntas de la izquierda. De manera similar, en el caso de P. cristatus (fig) el criterio binomial "conífera<0.5" nos indica los censos con presencia de coníferas se concentran en las puntas de la derecha.
Como ya se ha dicho, las hojas o puntas terminales de los árboles, informan de la abundancia o densidad media de todas las unidades muestrales que las componen. Tomando este hecho en combinación con los criterios de ramificación, se puede observar que, en A. caudatus, las hojas de la izquierda del árbol de abundancias del atlas de invernantes (y, por lo tanto, las unidades muestrales que las componen) tienen en general una menor abundancia de la especie que las hojas de la derecha (fig). Puesto que todas estas cumplen la condición de tener una baja proporción de suelo agrícola (primer criterio: agrícola>36%) podemos inducir que la variable agrícola afecta negativamente a la abundancia de A. caudatus (por lo que este criterio aparece con un signo "-" en la tabla 1). Por el contrario, el efecto de la variable usada en el primer criterio para abundancias de P. cristatus es positivo (fig): las unidades muestrales con mayor abundancia de esta especie se encuentran a la derecha, teniendo todas ellas más de un 11% de su superficie ocupada por masas de coníferas. Razonamientos equivalentes se han de seguir con respecto a los árboles relativos a los censos de densidades. Así, en ambas especies se observa en general una mayor densidad en las puntas terminales de la derecha: aquellos censos con presencia de arbolado caducifolio para el caso del Mito (fig), o los de mayor índice de estructura para el caso del Herrerillo Capuchino (fig).
El examen de las hojas terminales también ayuda a predecir cuáles son las mejores condiciones para las especies. Por ejemplo, la máxima abundancia predicha por los árboles desarrollados para los datos de abundancia del atlas es, en A. caudatus, de 82 aves/20 horas (fig). Esta máxima predicción se hace para las cuadrículas con menos del 36% de superficie agrícola y que estén al este de la coordenada UTM 0395152. No obstante, en otras zonas más occidentales, las condiciones más propicias para detectar una gran abundancia de mitos (78 aves /20 horas) se dan en áreas ricas en formaciones de matorral o menos complejas (criterio ‘estructura<3’) y con presencia de arbolado deciduo.
Los árboles de regresión podrían hacerse crecer, mediante la adición de criterios sucesivos, hasta que cada hoja correspondiera con una sola observación. Pero esto conduciría a un modelo excesivamente complejo, y poco o nada generalizable, por lo que en nuestro análisis hemos impuesto la condición de que cada una de las puntas terminales tuviera un mínimo de cinco unidades muestrales.
En el tipo de representación gráfica que hemos elegido la longitud de las ramas da idea de la importancia relativa de cada uno de los criterios que intevienen en ese árbol. De esta forma, las variables que hemos detectado como los principales determinantes de la abundancia invernal de A. caudatus en Madrid son los porcentajes de uso agrícola y de bosques de hoja caduca (deciduos) y un efecto geográfico determinado por la longitud (el mito es más abundante en las áreas orientales). Asimismo, para el caso de la densidad de esta especie, los principales determinantes (ramas más largas) son la presencia/ausencia de arbolado caducifolio y la complejidad estructural de la vegetación. Los tres principales determinantes de la distribución invernal de P. cristatus en Madrid parecen ser el porcentaje de masas de coníferas y, con notable menor importancia, los porcentajes de suelo ocupados por usos agrícolas y bosques deciduos. La densidad del Herrerillo Capuchino viene determinada básicamente por la estructura de la vegetación. En general, los criterios se ordenan de arriba abajo en función de su importancia pero esto no siempre ocurre así, como puede observarse en el gráfico para abundancias de A. caudatus. Además, una misma variable puede aparecer como criterio dicotómico en diferentes niveles a lo largo de un mismo árbol, y con distinta relevancia. Los tres criterios más importantes para cada especie se presentan en las tablas 1 (datos del atlas) y 4 (datos de los censos).
Si se tiene una nueva muestra (en nuestro caso un nuevo censo o una nueva cuadrícula del atlas) se puede predecir el valor de abundancia que cabría esperar que tuviera de acuerdo con el modelo del árbol de regresión. Para ello se procede paso a paso evaluando los distintos criterios desde el inicial hasta llegar a una de las puntas terminales (hay que recordar que cuando se cumple un criterio nos desplazamos hacia la izquierda y abajo en busca del siguiente criterio).
El grado de fiabilidad de los árboles de regresión puede evaluarse mediante el estadístico D2, conocido como el porcentaje de devianza explicada (D2% en las tablas 1 y 4), que toma valores entre 0 y 100%. Así, los árboles para A. caudatus y P. cristatus dan cuenta del 65.6 y 68%, respectivamente, de la información contenida en los datos de abundancia, y del 75.1 y 66.5%, respectivamente, para los de densidad. Los porcentajes de variabilidad restantes se deben a factores no incorporados a los modelos (e.g.: analíticos, como errores de muestreo o variables importantes que no hemos sabido identificar; biológicos, como relaciones interespecíficas; contingentes, como efectos históricos). Los árboles de regresión que hemos desarrollado aquí no alcanzan valores de D2 muy altos (media de 51% para los datos del atlas y 36% para los censos), aunque se encuentran dentro de los rangos típicos en estudios en ecología. Debe destacarse, no obstante, que puede considerarse un éxito alcanzar tales valores de D2, pues corresponden a la pretensión de resumir un fenómeno tan complejo como la invernada de aves en Madrid mediante unas pocas variables sencillas.
{kind=link}
{kind=link}
{kind=link}
{kind=link}